|
 Nguyễn Minh Nam Nguyễn Minh Nam
Trong 50 năm thuật ngữ gen được hiểu đồng nghĩa với những vùng RNA thông tin (mRNA) mã hóa cho bộ gen. Tuy nhiên các nghiên cứu gần đây đã chỉ ra rằng 90% bộ gen của sinh vật nhân thật được sao mã, trong đó chỉ một lượng rất nhỏ (khoảng 2-3%) các gen mã hóa protein, và một số RNA có chức năng trong giải mã protein (các RNA ribosome (rRNA), RNA vận chuyển (tRNA), phần lớn còn lại là các RNA không mã hóa protein (non-coding RNA, ncRNA) (1), (2), (3). Các ncRNA này thực hiện các vai trò trọng yếu trong tế bào như điều hòa phiên mã và sau phiêm mã, điều khiển phức hệ chỉnh sửa sợi nhiễm sắc (4), kích hoạt gen (2),... Ở bài này sẽ sơ lược về một số ncRNA và chức năng của chúng.
1. Long non-coding RNA
Là nhóm RNA rộng nhất, bao gồm toàn bộ các RNA không mã hóa protein có kích thước lớn hơn 200 nucleotide, gồm cả các mRNA giống ncRNA. Chúng có chức năng trong việc đóng xoắn của nhiễm sắc thể, sắp xếp cấu trúc các nội quan tế bào, telomere và điều hòa biểu hiện của các gen mã hóa protein bên cạnh (5). Ngược lại với sự điều hòa gen đã được thấy ở PTGS và TGS, ncRNA có thể cũng kích hoạt gen (hình 1) (2).
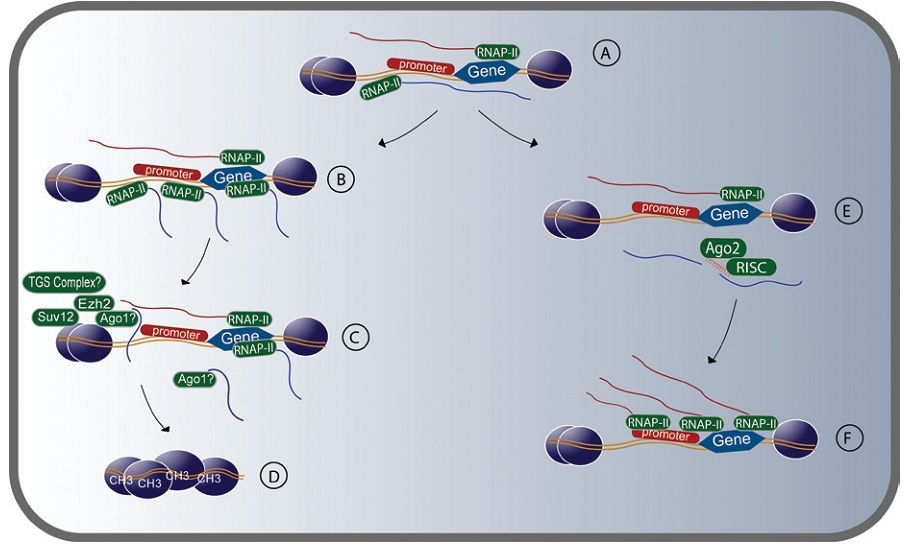
Hình 1. Chức năng điều hòa biểu hiện gen của các lncRNA ở tế bào động vật hữu nhũ. Ở bất kỳ thời điểm nào luôn có sự cân bằng giữa các bản sao sense và antisense của một gen được sao mã song hướng. (A) Sự biến động của tế bào có thể làm mất đi sự cân bằng này và ảnh hướng tới sự phiên mã của gen. Sự gia tăng phiên mã sợi antisense (B) dẫn đến sự gắn các protein điều chỉnh sợi nhiễm sắc vào gen promoter (C). Enzyme methyltransferases protein histon Ezh2 và Suvl2 điều hòa gen bởi các bản sao antisense (C). Sự im lặng bởi lncRNA liên quan đến các protein trong TGS như Ago-1, HDAC-1 (Histone deacetylase 1), và DNMT3A (DNA (cytosine-5)-methyltransferase 3A) (C). Sự gia tăng sự phiên mã các antisense đã hình thành sợi dị sắc và im lặng gen (D), tương tự như TGS. Ngoài ra, các bản sao antisense có thể bị cắt bởi siRNA theo cách phụ thuộc Ago-2 (E). Các bản sao antisense giảm làm tăng biểu hiện của gen (F) (2).
Khả năng nhận biết các trình tự bổ sung của các ncRNA cho phép các tương tác đặc hiệu cao tuân theo những bước điều hòa khác nhau trong quá trình sau sao mã của mRNA bao gồm sự nối, chỉnh sửa, vận chuyển, sao mã và thoái hóa (5).
Trên tế bào người, các sợi RNA antisense ức chế sự biểu hiện của các sợi sense bổ sung với chúng. Phương thức điều hòa này là do các cấu trúc thứ cấp được tạo ra từ các RNA sense và antisense hoặc do sự giải mã bị khóa bởi sự bắt cặp sense-antisense (6).
Các quan sát trên những gen ức chế ung thư đã phát hiện các ncRNA antisense là các yếu tố điều hòa sao mã. Sự quá biểu hiện của các ncRNA nào đó dẫn tới sự stable, epigenetic silencing các mRNA bổ sung tại promoter, trong khi sự mất các lncRNA antisense làm gia tăng sự biểu hiện của các mRNA bổ sung và vì thế gen được kích hoạt. ncRNA antisense có thể thay đổi cấu trúc vị trí sợi nhiễm sắc thông qua việc định vị tới gen đích, và tác động tới sự sao mã. Sự điều hòa sao mã trên S. cerevisiae, vi khuẩn và thực vật cũng được thực hiện bởi lnc RNA antisense. Như thế, nấm men, vi khuẩn, thực vật và con người cùng chung một cơ chế điều khiển sao mã bởi các lncRNA không phụ thuộc siRNA.
Các bản sao antisense không xuất hiện ngẫu nhiêm mà tập trung nhiều ở xung quanh các promoter của các vùng mã hóa và liên kết với các promoter có chứa các đảo CpG. Sự gia tăng sự methyl hóa DNA ở các promoter được điều hòa bởi một RNA antisense khi có sự suy giảm exosome RNA từ các tế bào.
Trong các gen liên quan tới ung thư, các gen mã hóa cho p21, E-cadherin, p15, p53, myc, vùng tương đồng yếu tố tăng trưởng biểu mô 1 (epidermal growth factor homology domain-1, tie-1) và PU.141 được điều hòa bởi các lncRNA. Một số gen có thể bị im lặng trên (ngoài) di truyền (epigenetic) do sự sản xuất bất điều hòa của lncRNA antisense. Giả thuyết này đặt ra câu hỏi rằng yếu tố nào điều hòa sự biểu hiện của các ncRNA antisense? Có thể giải thích là các miRNA điều khiển số lượng các vị trí gen được sao mã song hướng. Ví dụ trong trường hợp của E-cadherin, sự kết hợp của miRNA miR373 với E-cadherin ncRNA antisense làm tăng sự hiểu hiện của E-cadherin. Một lý giải khác là vai trò điều hòa bởi tiny RNA (tiRNA). Chúng chủ yếu định hướng sense và chỉ tập trung vào downstream của các vị trí khởi đầu sao mã (6).
Các ncRNA ức chế toàn bộ và trực tiếp RNA polymerase bằng sự bắp cặp các vị trí hoạt động và ngăn cản sự khởi đầu sao mã ngay từ những bước đầu tiên. Đồng thời cũng điều khiển sự phục hồi cấu trúc đặc biệt của các enzyme chỉnh sửa sợi nhiễm sắc (7).
Các lncRNA đóng một vai trò trong sự im lặng của các gen phát triển quan trọng và sự bất hoạt nhiễm sắc thể X trong động vật hữu nhũ. Sự bất hoạt X là sự đáp ứng sự im lặng của một choromosome X trên các cá thể cái để cân bằng biểu hiện gen với các cá thể đực. Hai lncRNA (RepA và Xist) có nhiệm vụ khởi đầu và duy trì sự bất hoạt X bằng cách hướng tới phức hợp chỉnh sửa sợi nhiễm sắc làm các nhiễm sắc thể bị im lặng, dẫn tới sự methyl hóa histone và hình thành sợi dị sắc (7).
2. Small non coding RNA
a. PIWI interacting RNA
PIWI-interacting RNA (piRNA) là các RNA dài 24-30nu, được tạo ra bởi cơ chế độc lập với Dicer và liên kết với protein PIWI, một nhóm phụ của protein Argonaute liên quan tới PIWI. PIWI protein chủ yếu có trong dòng tế bào mần. Hàng chục nghìn loại piRNA được tìm thấy trong động vật, zebrafish và ruồi giấm. Hầu hết các piRNA được tạo ra từ một số RNA tiền thân sợi đơn dài được mã hóa bởi các trình tư lặp lại trong bộ gen (8).
Tạo dòng các RNA nhỏ liên kết với các PIWI protein để nghiên cứu về piRNA trên mô hình động vật hữu nhũ đã được thực hiện năm 2006. Sau đó các piRNA đã được nghiên cứu trên các hệ thống mô hình khác. Các piRNA chủ yếu được sinh ra từ các transposon hoặc các trình tự lặp lại khác vì vậy nó còn có một tên gọi khác là rasiRNA (repeat-associated small interfering RNAs). Hiện nay các piRNA được chứng minh rõ ràng là được tạo ra từ cả các trình tự lặp lại và các trình tự DNA phức tạp khác và rasiRNA là một nhóm phụ của piRNA (8) (9).
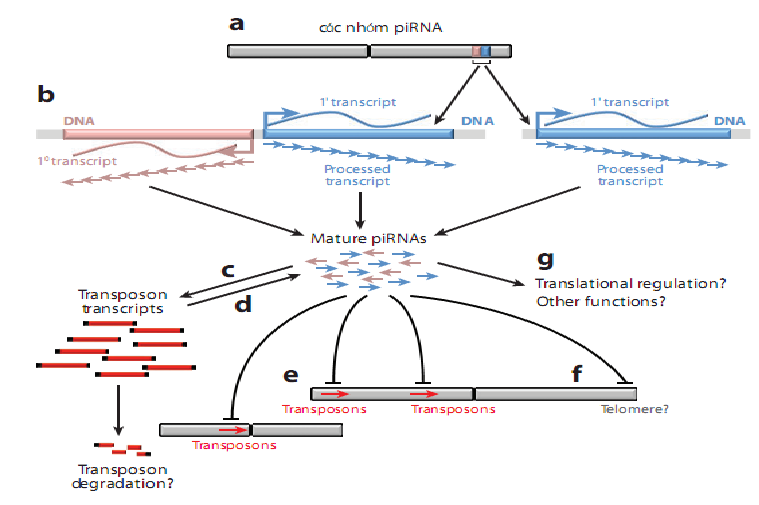
Hình 2: Sự hình thành và chức năng của các piRNA: piRNA được tạo thành tự các đoạn dài mã hóa bởi các bản sao sơ cấp. (a) Thông thường, các nhóm piRNA xuất hiện các bản sao song hướng (hiếm khi các bản sao sơ cấp được bắt nguồn từ một sợi đơn). (b) các nhóm được sao mã thành các bản sao dài sau đó đi vào quá trình loại bỏ các RNA dư và tạo thành các piRNA trưởng thành (cơ chế loại bỏ các RNA thứ cấp chưa được biết rõ). (c) piRNA được xem như một vector của sự điều hòa các mRNA transposon (tạo sự thoái hóa transposon). (d) Cùng lúc đó, sự tăng cường sau dịch mã hình thành nhiều piRNA trưởng thành hơn. (e) Có thể thông qua các piRNA dẫn tới đàn áp epigenetic của các vùng mã hóa transposon (mũi tên đỏ). (f) Các chức năng epigenetic khác của PIWI protein có thể tồn tại, trong ruồi giấm, PIWI protein có vai trò trong epigenetic và có thể trong điều khiển NST. (g) Bằng chứng cho thấy vai trò của piRNA trong điều khiển giải mã và các chức năng khác hiện diện nhưng chưa được kiểm tra nghiêm ngặt (8).
Ly trích các RNA tổng số từ chuột thí nghiệm thấy có chứa một lượng lớn các đoạn dài khoảng 30 nucleotide liên quan tới MIWI (protein PIWI của chuột). Bốn mươi piRNA bắt đầu được clone và từ 40 clone này đã xác định rằng các RNA nhỏ từ ít nhất một trong số các clone chứa piRNA.
Dựa vào phân đoạn gradient sucrose đã nhận thấy rằng piRNA hiện diện trong các mRNP, monosome, và polysome nhiều như nhau. Sự hiện diện của piRNA trong phân đoạn polysome cho thấy vai trò tiềm ẩn của piRNA trong việc điều khiển giải mã.
Các piRNA phân bố không đồng đều trong bộ gen, có hơn 96% được tập trung ở hàng trăm vị trí. Những nhóm này mang những đặc tính thú vị (hình 2).
piRNA có ảnh hưởng rộng lớn trong các tiến trình sinh học. Chúng giữ vai trò trọng yếu trong sự phát triển của các dòng mầm trên chuột, ruồi giấm và zebrafish và cần thiết cho sự sinh sản của cả ruồi giấm đực và cái. Tuy nhiên, các protein bao gồm sản xuất piRNA liên quan tới việc điều kiển biểu hiện gen trong các tế bào soma trong học tập và trí nhớ. (9).
b. Micro RNA
Một trong những bất ngờ lớn nhất đầu kỷ nguyên hậu gen (post-genome era) là việc phát hiện ra hàng loạt các gen mã hóa cho các microRNA. Các microRNA này được tìm thấy ở nhiều sinh vật khác nhau như Caenorhabditis elegans, Drosophila melanogaster, Arabidopsis Thaliana và người. Chúng có vai trò quan trọng trong sự tiến hóa của các sinh vật đa bào. (10).
MicroRNA là những nc RNA, tham gia vào sự điều hòa gen có chiều dài khoảng 22 nucleotide. Trên 3000 miRNA đã được tìm thấy trong nhiều loài, có vai trò trong việc phân hóa tế bào, sự phát triển, apoptosis, đề kháng virus và ung thư. Ứng dụng các phương pháp như real-time PCR và microassay kết hợp với những chương trình phần mềm để nghiên cứu sự biểu hiện và các chức năng mới của miRNA. Những kết quả này sẽ hé lộ vai trò điều hòa gen thực tế của miRNA trong từng khía cạnh cụ thể (11).
Nghiên cứu của Lee và cộng sự cho thấy sự điều hòa biểu hiện của các miRNA xảy ra ở nhiều mức độ bao gồm bước tách RNA dư (processing) và bước vận chuyển vào nhân. Các miRNA được biểu hiện đa nhân và được tách các RNA dư thông qua ít nhất 2 bước: (1) hình thành các pre-miRNA dài khoảng 70 nucleotide từ các bản sao dài hơn (được gọi là pri-miRNA); và (2) sự tách các đoạn RNA dư từ pre-miRNA thành miRNA (12).
c. siRNA
siRNA là các RNA ngắn có kích thước khoảng 20 tới 30 nu, được hình thành từ các RNA sợi đôi. Chúng liên quan tới một số protein họ Argomaute (Ago), chính protein này điều khiển chúng tới đích điều hòa (13).
Hiện tượng RNAi được khám phá đầu tiên trên giun tròn Caenorhabditis elegans do câm lặng gen bởi RNA sợi đôi (14). Theo sau đó những nghiên cứu của Guo và Kemphues đã phát hiện RNA sợi sense và antisen có hiệu quả ức chế biểu hiện gen như nhau (15). Sau đó Fire và cộng sự đã tập trung dùng antisenRNA để ức chế biểu hiện gen (14). Hiệu quả tác động của hỗn hợp sense và antisense RNA ít nhất gấp 10 lần so với chỉ là dùng sợi sense hay antisense (16). RNAi có thể bị kích động bằng việc tiêm các RNA sợi đôi vào tuyến sinh dục của C. elegans hoặc đưa vào bằng đường ăn thì cả dsRNA của C. elegans và dsRNA đưa vào đều biểu hiện.
Các siRNA được chuyển vào trong tế bào để nghiên cứu vai trò của protein trong việc đáp ứng phá hủy DNA và điều khiển chu kỳ tế bào, sự chuyển hóa chu kỳ tế bào cơ bản, tín hiệu và sự tái sắp xếp của chúng trong suốt quá trình phân bào, vận chuyển qua màng, sao mã và sự methyl hóa DNA. Những phân tử này có thể được dùng để phân biệt gen chủ với những gen khác (17).
Trong các tế bào người sự kích hoạt gen được thấy đầu tiên là các siRNA kích hoạt các promoter của E-cadherin, yếu tố tăng trưởng mạch nội mô (VEFG), và p21 làm tăng mức độ mRNA và protein của các gen này (6).
Trong in vivo, con đường siRNA đóng vai trò quan trọng trong việc hạn chế lây nhiễm virus vì nó làm bất ổn RNA được tạo ra trong chu kỳ sống của virus (9).
d. Small nuclear RNAs (snRNAs)
snRNA là các RNA không mã hóa, không chứa poly A và có chức năng trong nhân. Các snRNA có thể được chia thành 2 nhóm dựa trên những đồng yếu tố protein và các đặc tính trình tự chung (18).
e. Small nucleolar RNAs (snoRNAs)
snoRNA có kích thước khoảng 60 đến 300 nu, nhưng chỉ có những trình tự ngắn tham gia vào việc nhận biết trình tự đích thông qua các tương tác antisense. snoRNA điều khiển sự chỉnh sửa các vùng đặc hiệu của nucleotide trên các RNA đích. Có hai loại chỉnh sửa xảy ra, sự methyl hóa 2’-O-ribose và sự pseudouridylation (sự chuyển urudine thành pseudouridine), được điều khiển bởi các snoRNA được gọi là các snoRNA hộp C/D và hộp H/ACA (19).
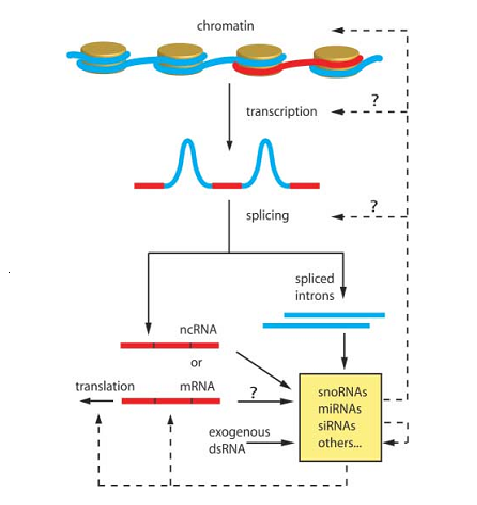
Hình 3. Hệ thống điều hòa bởi các sncRNA. Các RNA không mã hóa nhỏ điều hòa cấu trúc bộ gen và biểu hiện gen ở nhiều mức độ, miRNA, siRNA, snoRNA và các RNA nhỏ khác tham gia vào sự điều hòa giải mã, ổn định mRNA và cấu trúc sợi nhiễm sắc cũng như là tự điều hòa (nét đứt) và cũng có thể điều khiển giải mã và sự nối mRNA (dấu hỏi).
Chức năng chủ yếu của các RNA C/D và H/ACA là chỉnh sửa RNA ribosome và cắt các RNA dư trong nhân (18).
Tổng hợp về hệ thống điều hòa bởi các sRNA được tóm tắt ở hình 3.
3. Ứng dụng trong liệu pháp điều trị và chẩn đoán.
Các miRNA là những biomarker tốt trong chẩn đoán và tiên lượng ung thư. miRNA có tiềm năng lớn trong liệu pháp ung thư do những đặc điểm điều hòa gen. Các oligonucleotide kháng miRNA được thiết kế bổ sung với các miRNA của gen sinh ung thư có thể ức chế một cách đặc hiệu hoạt tính của miRNA trong các bướu. Nói cách khác, sự quá biểu hiện của miRNA có thể ức chế ung thư (20).
Dựa vào những thông tin về miRNA có thể phát hiện chính xác nguồn gốc của khối u và ung thư. Thực tế, chỉ cần khoảng 200 miRNA đặc trưng là có thể đủ để phân loại ung thư. Những khó khăn trong việc phát hiện sớm ưng thư ruột kết và một số ưng thư bí ẩn khác có thể được giải quyết bằng các dữ liệu miRNA thu được từ huyết thanh, huyết tương, nước bọt và một số mô khác của bệnh nhân. Giống như ung thư ruột kết, ung thư phổi và ung thư vú cũng liên kết chặt chẽ với các miRNA tương ứng. Điều này chứng tỏ vai trò tích cực của ncRNA trong chẩn đoán bệnh học.
Các miRNA không chỉ cung cấp một liệu pháp hứa hẹn trong điều trị ung thư mà còn hướng tới rất nhiều bệnh khác như sự nhiễm virus và các bệnh về tim mạch,… nơi mà các miRNA đóng vai trò như tác nhân điều hòa gen (20).
Sự phát triển những liệu pháp RNA không được thực hiện bởi vì sự bất ổn định của RNA (nhanh chóng bị phân hủy) in vivo. Tuy nhiên, DNA plasmid có thể được thiết kế để biểu hiện những đoạn RNA kẹp tóc ngắn (small hairpin RNA, short hairpin RNA - shRNA), giống như các miRNA ngoại lai. Liệu pháp gen dựa trên RNAi không có bản chất virus (non viral RNAi) kết hợp với công nghệ gen đích mới, sẽ gói DNA plasmid vào trong các thể liposome miễn dịch được pegylate (PILs) đặc hiệu cho thụ thể. Hướng này rất khả thi bởi hiệu quả đã đạt được. Knockdown 90% sự biểu hiện của gen đặc hiệu ung thư não. Tỉ lệ sống của chuột bị ung thư não đã được kéo dài gần 90% khi được tiêm dịch truyền chứa PILs chứa DNA plasmid mang shRNA trực tiếp kháng lại thụ thể yếu tố tăng trưởng biểu mô người. Liệu pháp gen dựa trên RNAi có thể được kết hợp với liệu pháp gen để thay thế những gen ức chế ung thư bị đột biến từ đó xây dựng chiến lược đa gen hướng tới liệu pháp gen trị ung thư. (21)
Liệu pháp gen dựa trên RNAi trong điều trị ung thư hiện nay sử dụng plasmid mang những đoạn shRNA đích tới các trình tự nucleotide trương đối ngắn (15-25nu), đặc hiệu cho những bản sao đột biến chỉ tìm thấy trong các tế bào ung thư (21).
Một số thuật ngữ:
Epigenetic: những sự thay đổi về kiểu hình bởi các cơ chế bên ngoài bộ gen. Những thay đổi như vậy có thể được duy trì thông qua quá trình phân bào, ví dụ như sự biệt hóa tế bào, hoặc chúng có thể được bảo tồn qua các thế hệ sau. Những thay đổi ngoài di truyền (epigenetic) bao gồm sự chỉnh sửa nhiễm sắc thể, ví dụ sự acetyl hóa protein histone, hoặc những thay thế hóa học trong chính DNA, ví dụ như sự methyl hóa DNA.
Sự bất hoạt nhiễm sắc thể X (X chromosome inactivation): là tiến trình mà trong đó 1 trong hai bản sao của nhiễm sắc thể X trên những cá thể cái bị bất hoạt. Sự bất hoạt xảy ra để mà những cá thể cái sinh ra cùng một lượng nhiễm sắc thể X bằng với cá thể đực (5).
PTGS (Post-transcriptional gene silencing): im lặng gen sau phiên mã: là kết quả của mRNA của một gene đặc biệt bị phá hủy hoặc bị khóa từ đó ngăn cản sự dịch mã của mRNA. Cơ chế chung của im lặng gen sau phiên mã là can thiệp RNA (RNA i).
TGS (Transcriptional gene silencing): im lặng gen trong phiên mã: là kết quả của những thay đổi histone, tạo ra môi trường dị nhiễm sắc xung quanh gen làm cho nó không thể tiếp cận với bộ máy phiên mã (RNA polymerase, các yếu tố phiên mã, vv.).
Promoter song hướng: là các trình tự promoter nằm ở giữa những cặp gen khác nhau và khởi đầu cho sự sao mã theo cả 2 chiều.
Antisense: sợi đối mã, có trình tự bổ sung với mRNA.
Sense: sợi mã hóa, có cùng trình tự với mRNA.
Ago: ARGONAUTE, họ protein kết hợp với các sRNA bao gồm siRNA và miRNA, có khối lượng phân tử 95 tới 100kDa. (22)
Dicer: là một endoribonulease trong họ RNase III, có chức năng cắt dsRNA và pre-miRNA thành những đoạn dsRNA ngắn gọi là siRNA, dài khoảng 20 – 25 nucleotide. Dicer xúc tác bước đầu tiên trong con đường RNAi và khởi đầu sự hình thành RNA-induced silencing complex.
Pegylated: Polyethylene glycol-coated: là quá trình gắn liên kết cộng hóa trị của các chuỗi polymer polyethylene glycol với các phân tử khác, thường là một loại thuốc hoặc protein trị liệu.
Histone deacetylases (HDAC) là lớp các enzym có chức năng loại bỏ các nhóm acetyl từ lysine trên histone.
DNA (cytosine-5)-methyltransferase 3A:
CpG site: là những vùng DNA chứa nucleotide cytosine (C) kế guanine (G). CpG là dạng viết tắt của "—C—phosphate—G—", có nghĩa là cytosine và guanine được phân cách và nối với nhau bởi 1 phân tử phosphate trên DNA. Ký hiệu CpG được dùng để phân biệt trình tự này với cặp CG của cytosine và guanine.
Đảo CpG: là vùng DNA xuất hiện nhiều CpG. Nhiều gen của động vật hữu nhũ chứa các đảo CpG liên kết với điểm khởi đầu của gen. Vì thế, sự hiện diện của các đảo CpG được dùng để giúp tiên đoán và giải thích các gen.
1. Costa, F. F. (2010) Non-coding RNAs: Meet thy masters, Bioessays 32, 599-608.
2. Turner, A. M., and Morris, K. V. (2010) Controlling transcription with noncoding RNAs in mammalian cells, Biotechniques 48, ix-xvi.
3. Ambros, V. (2001) microRNAs: tiny regulators with great potential, Cell 107, 823-826.
4. Taft, R. J., Pang, K. C., Mercer, T. R., Dinger, M., and Mattick, J. S. (2010) Non-coding RNAs: regulators of disease, J Pathol 220, 126-139.
5. Mercer, T. R., Dinger, M. E., and Mattick, J. S. (2009) Long non-coding RNAs: insights into functions, Nat Rev Genet 10, 155-159.
6. Morris, K. V., and Vogt, P. K. (2010) Long antisense non-coding RNAs and their role in transcription and oncogenesis, Cell Cycle 9.
7. De Lucia, F., and Dean, C. (2010) Long non-coding RNAs and chromatin regulation, Curr Opin Plant Biol.
8. Thomson, T., and Lin, H. (2009) The biogenesis and function of PIWI proteins and piRNAs: progress and prospect, Annu Rev Cell Dev Biol 25, 355-376.
9. Klattenhoff, C., and Theurkauf, W. (2008) Biogenesis and germline functions of piRNAs, Development 135, 3-9.
10. Krol, J., and Krzyzosiak, W. J. (2004) Structural aspects of microRNA biogenesis, IUBMB Life 56, 95-100.
11. Wang, Y., Stricker, H. M., Gou, D., and Liu, L. (2007) MicroRNA: past and present, Front Biosci 12, 2316-2329.
12. Lee, Y., Jeon, K., Lee, J. T., Kim, S., and Kim, V. N. (2002) MicroRNA maturation: stepwise processing and subcellular localization, EMBO J 21, 4663-4670.
13. Ghildiyal, M., and Zamore, P. D. (2009) Small silencing RNAs: an expanding universe, Nat Rev Genet 10, 94-108.
14. Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., and Mello, C. C. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans, Nature 391, 806-811.
15. Guo, S., and Kemphues, K. J. (1995) par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed, Cell 81, 611-620.
16. Timmons, L., and Fire, A. (1998) Specific interference by ingested dsRNA, Nature 395, 854.
17. Agrawal, N., Dasaradhi, P. V., Mohmmed, A., Malhotra, P., Bhatnagar, R. K., and Mukherjee, S. K. (2003) RNA interference: biology, mechanism, and applications, Microbiol Mol Biol Rev 67, 657-685.
18. Matera, A. G., Terns, R. M., and Terns, M. P. (2007) Non-coding RNAs: lessons from the small nuclear and small nucleolar RNAs, Nat Rev Mol Cell Biol 8, 209-220.
19. Mattick, J. S., and Makunin, I. V. (2005) Small regulatory RNAs in mammals, Hum Mol Genet 14 Spec No 1, R121-132.
20. Angaji, S. A., Hedayati, S. S., Poor, R. H., Madani, S., Poor, S. S., and Panahi, S. (2010) Application of RNA interference in treating human diseases, J Genet 89, 527-537.
21. Pardridge, W. M. (2004) Intravenous, non-viral RNAi gene therapy of brain cancer, Expert Opin Biol Ther 4, 1103-1113.
22. Mourelatos, Z. (2007) Small regulatory RNAs: biogenesis & functions, Nucleic Acids Symp Ser (Oxf), 105.
Số lần xem trang : 15800
Nhập ngày : 23-05-2011
Điều chỉnh lần cuối : Ý kiến của bạn về bài viết này
 Sản xuất taxol từ nấm nội sinh thực vật(14-07-2011) Sản xuất taxol từ nấm nội sinh thực vật(14-07-2011)
Tam giác Nano bạc bọc chitosan, tiền năng trong điều trị ung thư(27-06-2011) Tinh dầu cá và tinh dầu lưu ly có thể giảm viêm và loãng xương(18-06-2011) Tác dụng chống ung thư gan của Cannabinoids từ cây cần sa (17-06-2011) Tạo tế bào thần kinh từ tế bào thường(08-06-2011) Tầm nhìn phát triển vaccine chống HIV, lao và sốt rét tới năm 2020(05-06-2011) Cách pha primer(01-06-2009) Bệnh vàng gân lá trên mía và sugarcane yellow leaf virus(05-03-2009) Một số bệnh do virus trên cây mía(05-03-2009) Trang kế tiếp ...
|




 Đang xem 4
Đang xem 4
 In trang này
In trang này Lên đầu trang
Lên đầu trang Gởi ý kiến
Gởi ý kiến